第一章 绪论
P1生物反应工程 是一门以生物学、化学、工程学、计算机与信息技术等多学科为基础,研究生物反应过程中带有共性的工程技术问题的交叉学科。
P1生物反应过程与化学反应过程本质的区别是有生物催化剂参与反应。
P1生物反应的特点:1)反应在温和的条件下进行;2)反应速率比化学反应过程慢很多;3)反应的复杂性有时难以预计。
P2生物反应过程研究的目的是提供适宜的动力学速率方程,以描述微生物(或酶、动植物等)反应体系,确定这些方程在设计方面的用途,规划实验室的实验、决定动力学方程所需的速率常数。
P2一般生物反应过程可以分为上游、中游和下游三个阶段,上游主要指原料处理与培养基制备,包括种子的制备,中游是利用生物反应获得目的产物的过程,下游是指目的产物的提取与精制。
P3 1971年,英国学者巴特金逊(Batkinson)首次提出生化反应工程这一术语。
P4生物反应工程的主要内容可分为生物反应动力学、生物反应器和生物反应过程的放大与缩小。
P5生物反应工程研究的是工业规模的生物反应。
第二章
P8生物反应过程中,各种物料具有不同的物理量和参变数,这些物理量和参变数是由几个独立的基本量组成,这种基本量称为因次。
P9基本量的大小以测量单位来表示,测量单位采用国际单位制(SI)。
P9单位质量物体的体积称为比容,比容是密度的倒数。
P9气体的体积与温度成正比,与压强成反比,因此气体的密度完全取决于温度和压强。
P10流体垂直作用于单位面积上的力称为压力强度,简称压强,但习惯上多称为压力。
P10压力大小通常用两种不同的基准表示:一种是以绝对真空为基准,称为绝对压强,简称绝压;一种是以当地大气压为基准,通常由压力表或真空表测出,称为表压或真空度。
P11单位时间内流过管道任一横截面的流体量称为流量。如流体用体积来度量,称为体积流量。若流体量用质量来计算,则称为质量流量。
P11流速是单位时间内流体在流动方向上所流经的距离,也就是流体的体积流量除以管道截面积。
P12粘度的物理意义是促使流体流动产生单位速度梯度时剪应力的大小。
P12流体的粘度还可用粘度μ与密度ρ的比值来表示,这个比值称为运动粘度。
P13温度对流体粘度的影响很大,温度升高时流体粘度剧烈下降,相反气体的粘度则随气温上升而增大。
P13雷诺准数与流体的流速、流体的密度、粘度及管道的直径有关系。
P13当Re<2000时,属于层流,当Re>4000时,属于湍流。
P13雷诺准数是一个无量纲的数群,故其值不会因采用的单位值不同而改变。
P13几个物理量组合而成的无量纲数群称为准数。
P14 用如下参数来描述剪切力(液体中)大小:(1)桨尖线速度;(2)平均剪切速率;(3)湍流漩涡长度。
P15物料衡算是依据质量守恒定律,对于选定的衡算系统,进入与离开这一过程系统的目的物料的质量之差,等于该过程系统中目的物料的消耗量和累计量之和。
P16物系的平衡关系是自然界广泛存在的现象。平衡关系可用来判断过程能否进行,以及进行的方向和能达到的限度。
P17速率是变化的快慢程度,有反应速率和传质速率两方面含义。针对具体的生物反应过程,其反应速率为单位时间、单位反应体积生成的产物量。
P17传质速率是单位面积上单位时间的传递量,与推动力成正比,与阻力成反比。
P17针对一个多步的生物反应过程,如果其中的一个步骤的反应速率低于其他步骤的,这一步骤将成为限制性步骤或者控制步骤。如果能够提高这一限制性步骤的反应速率,将使整个反应速率得到提高。
第三章 酶促反应动力学
P19酶促反应的特征:优点:常温、常压、中性范围(个别除外)下进行反应;与一些化学反应相比,省能且效率较高;专一性好;反应体系较简单,反应过程的最适条件易于控制等。不足:多限于一步或几步较简单的生化反应过程;一般周期较长。
P20酶是具有生物催化功能的生物大分子,按照分子中起催化作用的主要组分的不同,自然界中天然存在的酶可以分为蛋白类酶和核酸类酶两大类别。
P20蛋白类酶可分为六大类:氧化还原酶类、转移酶类、水解酶、裂合(解)酶类、异构酶、连接(或合成)酶。
P21酶的催化共性:a降低反应的活化能;b加快反应速率;c不能改变反应的平衡常数,只能加快反应达到的平衡的速度。
P21酶的生物催化特性:1)酶的专一性(一种酶只能催化一种或一类底物);2)高催化效率(反应速率大大加快);3)反应条件温和(几乎都在常温、常压、中性pH);4)酶活性受到调节(可被其他因素调节)。
P21酶活力的调节方式主要有酶浓度的调节、激素调节、共价修饰调节、限制性蛋白水解作用的调节、激活剂和抑制剂调节、前馈和反馈调节、金属离子和其他小分子化合物的调节、产能反应和需能反应的调节等。
P22酶失活的原因可以概括为4类:1)酶活性中心特定氨基酸残基被化学修饰;2)外部环境的影响,酶活性中心出现空间障碍,使其不能与底物相结合;3)酶的高级结构发生变化,相对而言是一种宏观变化;4)多肽链的断裂,可以说是一种“激烈的分解作用”。
P22确保酶活力稳定的稳定化方法及原因:低温(难以被化学物质或蛋白酶等破坏);添加盐类(如加入高浓度硫铵后,增强酶高级结构的稳定性);添加底物(保护酶的活性中心,有利于酶分子高级结构的稳定);添加有机溶剂(如加入丙酮,但机制不清楚);化学修饰(稳定酶的高级结构,保护酶的活性中心);加入强变性剂(只要保留一级结构, 仍可再生);加入蛋白质(加入清蛋白等用于保护在稀薄状态下易发生变性失活的酶类);结晶化(有利于高级结构的稳定);固定化(防止或减少蛋白酶的作用)。
P23固定化酶不仅指通过化学或者物理方法结合于固体支持物上或凝胶内的酶,还指那些被超滤膜或中孔纤维膜物理固定于有限空间的溶液酶。
P23酶的使用过程中要求酶的制备应具有以下特点:1)生产成本低;2)不受季节地区条件的限制;3)制造周期短;4)便于大规模生产等。
P23固定化后酶性质发生变化,主要表现在以下的4个方面:1)底物专一性的改变;2)稳定性增强;3)最适ph和最适温度的变化;4)动力学常数的变化。
P24酶的固定化方法可分为:1)吸附法;2)载体结合法;3)交联法;4)包埋法。各个方法的定义。
P25单底物均相反应系统中,如果酶促反应速率与底物浓度无关,此时为零级反应;当反应速率与底物浓度的一次方成正比,称为一级反应。
P27单底物的不可逆酶促反应是最简单的酶促反应。水解酶、异构酶及多数裂解酶的催化反应均属此类。
P27对于酶促反应,Henri提出活性中间复合体学说:认为酶促反应至少包括两步,首先是酶E与底物S相结合形成中间复合体ES,然后复合体分解生成产物P,并释放出酶E。
P28基于米氏方程的三点假说提出两种模型,即快速平衡学说和稳态法。如今常说的米氏方程是由稳态法推导的结果。
P29米氏方程中最为重要的动力学参数是Km和rp,max。使用较多的求以上两参数的方法是Lineweaver Burk法,又称双倒数作图法。另外误差较小的方法是Eadie-Hofstee法。
P29Lineweaver Burk法是将米氏方程取倒数,公式为

以1/rp对1/[s]作图得一直线,该直线斜率为km/rp,max,直线与纵轴交于1/rp,max。
P29米氏方程中求取动力学参数是Km和rp,max的Lineweaver Burk法的主要问题是当底物浓度[S]很低时,相应1/rp值也很小,取其倒数则使数据误差进一步放大,因此在底物浓度较低时,不适合采用此法。
P33实际酶促反应中,反应时间与底物转化率关系公式3-30

P35酶分子上有许多酸性或碱性的氨基酸侧链基团,这些基团处于适当解离状态时,酶才能发挥其活性。通常酶只有在一定pH范围内才具有活性。
P36在一定范围内,酶促反应速率随温度升高而升高;超过某一温度限制后,酶的活性随着温度升高而降低。
P38酶促反应速率降低原因:失活作用和抑制作用。
P38酶的不可逆抑制可能通过加入螯合剂EDTA或柠檬酸盐来解除,但不能通过透析、超滤等物理方法进行解除。
P38酶的可逆抑制剂与酶之间以非共价键结合,可以通过透析、超滤等物理方法进行解除。
P39有竞争性抑制剂存在时的动力学方程3-46

P39竞争性抑制改变了米氏常数,当[I]增加或ki减少,都导致米氏常数增大,因而降低了底物的反应速率,可以通过提高底物浓度来消除竞争性抑制。
P39非竞争性抑制剂与酶结合的位点不是酶的活性部位,而是酶的非活性部位。
P39非竞争性抑制剂存在时的动力学方程3-49

P39非竞争性抑制的最终结果是降低了rp,max,因此高浓度底物不能消除非竞争性抑制作用。
P40反竞争性抑制剂不能与游离酶结合,只能和酶-底物形成的复合物结合,形成酶-底物-抑制剂的复合物,导致不能生成产物,使酶催化反应速率下降。
P40反竞争性抑制的结果是rmax和km都降低,最终使酶催化反应速率降低。
P40对于某些酶促反应,当底物浓度较高时,反应速率呈下降趋势,称为底物抑制。
P41底物抑制建立的反应动力学方程3-54

P43影响固定化酶促反应的主要因素:分子构象的改变、位阻效应、微扰效应、分配效应、扩散限制。
P46Da准数的物理意义是最大反应速率与最大传质速率之比。
P46当Da准数愈小,固定化酶表面浓度[S]s愈接近于主体浓度[S],表面最大传质速率愈大于最大反应速率,过程为反应控制。
P46当Da准数愈大,固定化酶表面浓度[S]s愈小,愈接近于零,表明最大反应速率愈大于最大传质速率,过程为传质控制。
P47固定化酶促反应中,外部扩散过程的效率因子是指由外部扩散影响时的实际反应速率和无外部扩散影响时的固定化酶外表面处的反应速率之比。
P47Da准数是决定效率因子ηout和比浓度C*的唯一参数,是表征传质过程对反应速率影响的基本准数。
P47对于具有大量内孔的球形固定化酶颗粒,其内部是酶促反应的主要场所。底物通过孔口向内扩散,达到不同深度,产物沿反方向从内部向孔口外部扩散。
P47对于内部扩散过程的效率因子ηin可定义为单位时间内实际反应效率与按颗粒外表面底物浓度计算而得到的反应效率之比。
P48西勒准数的物理意义是表面浓度下的反应速率与内扩散速率之比。
P48普遍化的西勒准数的定义式为:

P49对于球形固定化酶颗粒的内部扩散效率因子,为颗粒的实际有效反应速率和颗粒内无浓度梯度时的反应速率之比。
P50对于球形固定化酶颗粒的内部扩散过程中,β对ηin影响不大,影响ηin的主要参数是西勒准数Φ。如果Φ<<0.3,则ηin不随Φ变化,近似等于1,也就是说没有内部传质阻力,若Φ>>0.3,则ηin<1,反应为内扩散所限制。
P50为提高固定化酶内扩散效率,应设法减小Φ。减小Φ的措施主要是适当降低固定化酶颗粒粒径。
P51酶的失活作用是指物理或者化学的因素部分或全部破坏了酶的三维结构引起酶的变性,导致酶一部分或者全部丧失活性。
P51酶在保存过程中,由于时间原因而使其失活该失活类型称为酶的静态稳定性。
P51如果酶在参与反应过程中发生了失活,则该失活过程被称为酶的操作稳定性。
P52多数酶的热失活反应服从一步失活模型,其中的Kd为一步失活常数,它的倒数称为时间常数tc,[E]t降至[E]0值1/2的时间称为半衰期t1/2,它们之间的关系式为

P52未反应时酶的热失活动力学中,多步失活模型分为多步串联失活模型和同步失活模型。
P52同步失活模型认为全部酶分子可以划为若干个热稳定性不同的组分,每个组分又均符合一级失活模型,全部酶[E]0中残存活性酶的比例Φ(t)为

其中Xi为失活速率常数为ki的酶组分的百分率。
P52温度对酶失活速率的影响体现在失活速率常数上。对于一级失活模型,有如下方程:

其中Ad为指前因子,Ed为失活反应活化能。
如果同时温度和时间对失活的影响,有下列公式:

P53反应中酶的热失活动力学中,当时间一定,随着温度的升高,反应速率增大,转化速率提高。当温度升高到某一个值,由于酶的热失活速率加快,当其快于酶促反应速率上升的速度时,酶的总反应速率下降,最终降为零。反应中酶失活模型的失活动力学方程可表示为:

P56双底物酶促反应中,有序连续机制的反应速率方程是:

第四章 微生物反应动力学
P82微生物菌体的80%左右是水分。湿菌体所含水分是指菌体在100℃前后干燥直到恒重时减少的量。除去水分的菌体称为干菌体。
P83细菌以分裂方式进行的繁殖。在适宜的生长条件下,某些细菌的世代时间可达10~20min。然而,比较典型的世代时间为40~60min。
P83酵母菌的生长方式有出芽繁殖、裂殖和芽裂(如同菌丝生长)三种。在最适条件下,酵母在45min内就可以分裂,比较典型的分裂时间为90~120min。
P83霉菌的生长特性是菌丝伸长和分枝。
P84相较于化学反应,微生物反应的缺点有:1、副产物的产生不可避免。2、影响微生物反应的因素多实际控制有难度;3、原料是农副产品,受价格变动影响大;4、产前准备工作量大,相对化学反应器而言,反应器效率低。对于好氧反应,需氧,故增加了生产成本,且氧的利用率不高;5、废水有较高BOD值。
P85碳源是指可构成微生物细胞和代谢产物中碳架来源的营养物质。
P85氮源主要是提供合成原生质和细胞其它结构的原料,一般不提供能量。
P85无机元素的主要功能是:构成细胞的组成成分;作为酶的组成成分;维持酶的作用;调节细胞渗透压、氢离子浓度和氧化还原电位等。
P85工业生产中,常利用玉米浆等作生长因素的供应量。
P86当溶解氧的浓度较低时,可以使用以培养基的氧化还原电位Eh来定量表示厌氧程度的方法。
P86好氧性微生物在Eh值为+0.1伏以上均可生长,以Eh等于+0.3~+0.4伏时为适。厌氧微生物只能在Eh值小于+0.1伏以下生长。兼性厌氧微生物在+0.1伏以上或以下均能生长。
P86细菌要求水活度(湿料饱和蒸汽压/相同温度下纯水饱和蒸汽压)在0.90~0.99之间;大多数酵母菌的为0.80~0.90;真菌及少数酵母菌要求在0.60~0.70。
P88可通过测定O2的消耗速率与CO2的生成速率来确定好氧培养中微生物生物代谢机能的重要指标之一的呼吸商。
P88呼吸商的定义式:
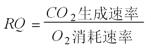
P90消耗1g基质生成细胞的克数称为细胞得率或称生长得率Yx/s。
P90同一菌种,同一培养基,好氧培养的Yx/s比厌氧培养的大得多。
P91当基质为碳源,无论是好氧培养还是厌氧培养,碳源的一部分被同化为细胞的 组成部分,其余部分被异化分解为CO2和代谢产物。
P91和碳元素相关的细胞得率Yc的计算公式:

P91Yave-是以有效电子为基准的细胞得率;Ykj为以总有效能为基准的细胞得率;YATP为以生成的ATP为基准的细胞得率。
P94微生物反应是放热反应。储存于碳源中的能源,在好氧反应中有40%~50%的能量转化为ATP,供微生物的生长、代谢之需,其余的能量作为热量被排放。
P94微生物反应中的反应热,也称代谢热。
P94采用复合培养基时,营养组分通过分解代谢,在生成能量(ADP→ATP)的同时,生成产物。另一方面,培养基中的组分通过同化代谢在合成微生物细胞的同时利用了能量(ATP→ADP)。这就是说能量可以从呼吸(如糖在氧存在下氧化、分解为CO2和H2O)和发酵(厌氧进程中糖分解为中间代谢物和CO2)获得。
P95一般能量偶联型生长(即生成ATP的分解代谢途径是细胞生长的限制因素),Ykj值较大;能量非偶联型生长Ykj值较小,能量利用效率比较差。
P99平衡生长条件下微生物细胞的生长速率rx的定义式为:
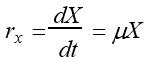 ;式中X为微生物的浓度,μ为微生物的比生长速率。
;式中X为微生物的浓度,μ为微生物的比生长速率。P99μ与倍增时间td的关系为
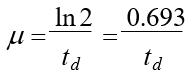
P100 微生物生长动力学的数学模型,分为下面几类:
(a)确定论的非结构模型,是一种理想状况,不考虑细胞内部结构,每个细胞之间无差别,细胞群体作为一种溶质。
(b)确定论的结构模型,每个细胞之间无差别,细胞内部有多个组分存在。
(c)概率论的非结构模型,不考虑细胞内部结构,每个细胞之间有差别。
(d)概率论的结构模型,细胞内部结构有差别,每个细胞之间也有差别。
一般人们提到的微生物生长动力学模型指的是确定论模型。
P101目前最易使用的模型是确定论的非结构模型。
P101Monod方程的基本形式:
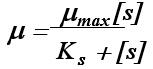
P106基质的消耗速率被菌体量除称之为基质的比消耗速率以希腊字母γ 来表示
P107Qo2为氧的比消耗速率,常称为比呼吸速率
P109代谢产物的生成速率中,当以体积为基准时,称为代谢产物的生成速率,记为rp;当以单位重量为基准时,称为产物的比生成速率,记为π
P109Gaden根据产物生成速率与细胞生成速率之间的关系,将其分成三种类型:相关模型;部分相关模型;非相关模型
第五章
P116好氧培养可采用以下几种方法:(1)液体表面培养(如使用浅盘);(2)通风固态发酵;(3)通氧深层培养。
P116就操作方式而言,深层培养可分为:分批式操作、半分批式操作、反复分批式操作、反复半分批式操作、连续式操作。
P116反复分批式操作是指分批操作完成后,部分取出反应物料,剩余部分作下一批培养的种子,重新加入一定量的基质,再按照分批式操作方式,反复进行。
P116半分批式操作,又称流加操作,反应过程中将特定的限制性基质按照一定要求加入到反应器内,以控制限制性基质保持一定。
P117反复半分批式操作是指流加操作完成后,不全部取出反应物料,剩余部分重新加入一定量的基质,再按照流加操作方式进行,反复进行。
P117连续式操作是指在分批式操作进行到一定阶段,一方面将基质连续不断地加入反应器内,另一方面又把反应物料连续不断的取出,使反应条件(如反应液体积等)不随时间变化的操作方式。
P117活性污泥法处理废水、固定化微生物反应等多采用连续式操作。
P118分批式操作的不足:反应器的非生产周期较长;由于频繁杀菌,易使检测装置损伤;由于每次培养均要接种,增加了生产成本;需要非稳定过程控制费用;人员操作加大了污染的危险。
P118流加式操作的不足:有反应器的非生产周期;需要较高的劳动力(需要控制和高价的检测装置);人员的操作加大了污染的危险;由于频繁杀菌,易使检测装置损伤。
P118分批式培养过程中,微生物的生长可分为:1)迟缓期2)加速区3)对数生长期4)减速期5)静止期6)衰退期
P120分批式培养过程的状态方程式(环境过程的状态方程式)可表示为:

氧:
CO2:
F为惰性气体流速,V为反应液总容积,Pall为气体总压力,(Po2)out 为排气中氧的分压,(Po2)in为进气体中氧的分压,(Pco2)in为进气体中CO2的分压,(Pco2)out为排气中CO2的分压。
P121可以认为分批培养过程中的动态特性取决于基质与微生物浓度(接种量)及微生物反应的诸比速率的初始值,因此,支配分批式培养统的主要因素是基质与微生物的浓度的初始值。
P123反复分批操作示意图
菌体量的衡算式为:
产物浓度的衡算为:
滤液取出率为:
产物的生产能力:
由上式可知,为提高产物生产能力,可采取提高α'+β'或减少 tRB
P124流加操作的优点是能够任意控制反应液中基质浓度。流加操作的要点是控制基质浓度。
P124从流加方式看,流加操作可分为无反馈控制流加操作与反馈控制流加操作。前者包括定流量流加、指数流加和最优流加量流加培养等。后者分间接控制、直接控制、定值控制和智能控制等流加操作。
P125流加操作时,特定基质加入到反应器后,反应液体积就会发生变化,状态变量的估算与质量衡算以总量的变化来进行。这时μ、γ和π的可定义如下:
式中,V为反应液体积,F是体积流量,[S]in是流加液中的基质浓度,F[S]in为基质的质量流量。
P125无反馈控制的流加操作,限制性基质的平衡式:
P126定流量流加操作是指基质的流加速度保持一定的流加操作。
P126定流量流加操作,时间t时,菌体的恒算式
P126定流量流加操作,最大特点是微生物进行线型生长。
P126指数流加操作是通过采用随时间呈指数性变化的方式流加基质,维持微生物菌体的对数生长的操作方法称为指数流加操作。此时,以满足μ等于定值为基础,流加基质,由Monod方程可获得[S]=常数。
P127微生物每次培养都可能有微妙的变化,因此,无反馈控制的流加操作适用范围很窄。
P127反馈控制的流加式操作可分为间接(与过程密切相关的、可以测定的参数为控制指标,如pH、DO、Qco2)和直接(连续或间接地测定培养液中流加的基质浓度,以此作为控制指标)两类。
P127反馈控制的流加式操作根据流加基质浓度的情况,可分为保持一定浓度值(定值控制)和浓度随时间变化(程序控制)两类控制方法。
P130连续操作有两大类型,即连续操作搅拌式反应器CSTR(continuous stirred tank reactor)型和连续操作活塞式反应器CPFR(continuous plug flow tulular reactor)型。
P130CSTR型连续操作,大致可分为两种。一是恒化器法(chemostat),二是恒浊器法(turbidstat)。
P130恒化器法是指在连续培养过程中,基质流加速度恒定,以调节微生物细胞的生长速率与恒定流量相适应的方法。
P130恒浊器法是指预先规定细胞浓度,通过基质流量控制,以适应细胞的既定浓度的方法。
P130单级CSTR培养系统中,流入液中仅一种成分为微生物生长的限制性因子,其他成分在不发生抑制的条件下充分存在。
第六章 动植物细胞培养
P165动植物细胞培养技术是一项将动植物的组织、 器官或细胞在适当的培养基中进行离体培养的技术。
P165由于动物细胞体外培养具有明显的表达产物的优点,为传统微生物发酵所无法取代,因此,生物技术中许多有价值的生物制品,需要借助于动物细胞培养而获得。
P165 1950年前后,Enders及其同事发表了第一篇关于在培养细胞中生长病毒的报告,开拓了以动物病毒为研究对象的新领域。
P166随着基因工程技术的发展,人们逐渐认识到有许多基因产物不能在原核细胞内表达,它们需要经过真核细胞所特有的翻译后修饰,以及正确的切割、折叠后,才能形成与自然分子一样的功能和抗原性。使得动物细胞一跃成为一种重要的宿主细胞,用以生成多种生物制品,例如病毒疫苗、淋巴因子(干抗素和白细胞介素等)、单克隆抗体、红细胞生成素、纤维蛋白溶酶原激活剂、第八因子、肿瘤坏死因子、上皮生长因子和过氧化物歧化酶等。
P166动物细胞培养与微生物培养的不同点:动物细胞无细胞壁,机械强度低,适应环境能力差;生长速度缓慢,易受微生物污染,培养时需要抗生素;大多数哺乳动物需附着在固体或半固体的表面生长;对营养要求严格;大规模培养时,不可简单地套用微生物培养的经验。
P168动物细胞生长模型:Monod方程、 Logistic方程、 底物和产物抑制模型。
P170动物细胞培养方法有两种类型:非贴壁依赖性细胞的培养和贴壁依赖性细胞的培养。
P170适用于动物细胞的还有一种连续式操作方法——灌流培养法。这种方法是把细胞置于某种封闭系统中,连续注入新鲜培养基的同时,连续等量排出用过的培养基,但不排出悬浮的细胞。采用这种方法后,从产物开始分泌时起,便可用快速纯化方法不断的从流出培养基中收集目的产物。虽然灌流培养系统具有如上优点,但也存在培养基消耗量比一般悬浮培养高几倍,工作过程复杂,培养物易受污染和培养细胞不稳定等缺点。
P171灌流培养系统可分为3类:(1)微小均质系统,如灌流悬浮细胞培养,微载体系统。(2)巨大均质系统,如把细胞包埋在微小球或凝胶中。(3)非均质系统,如边灌流边用过滤膜将细胞加以固定。
P173 1937年前后,法国和美国科学家分别成功的进行了胡萝卜和烟草的组织培养。
P173 1966年Klein指出,能利用植物细胞培养物代替收集或栽培植株来生产生化制品。
P173 20世纪90年代以来,伴随着紫杉醇的研究成功并应用于临床,植物细胞培养及其次级代谢产物的研究进入了新的发展时期,尤其是诱导子、前体饲喂、两相法培养、质体转化、毛状根和冠瘿瘤组织培养等新技术和新方法的发现和发展,更显示了植物细胞培养工程制药的广阔发展前景。
P176 微生物反应中,细胞的生长速率与限制性基质间的关系常用Monod方程来描述。同理,Monod方程也可以用于表达某些碳源与动植物细胞生长速率间的关系。
P176由于动植物细胞的培养过程是好氧过程,因此,许多科技工作者研究了与细胞呼吸速率相关的因素。例如,人参细胞培养中,添加椰子汁后细胞的生长速率加快,耗氧量较不添加椰子汁时明显增大。
P176胞内物质与生长模型:微生物培养中,把细胞内RNA含量作为描述其生长速率的指标。且随微生物比生长速率的增加,RNA含量增大。吉田等在进行人参细胞培养中发现,在对数生长期,细胞的RNA含量显著增大。
P177植物细胞的培养方式:根据所处状态的不同,分为悬浮培养与固定化植物细胞培养法,根据操作方式的不同,又分为分批式、反复分批式和连续式培养3种。
P177分批培养中,植物细胞的生长曲线一般呈 S型,可明显观察出诱导期、对数生长期、减速期、静止期和衰亡期。但是与细菌和酵母菌的培养相比,即使是生长速率快的烟草细胞,分批培养时间(一个周期)也要 6天以上,生长缓慢。
P177植物细胞培养中,藤田开发的二步培养法是分批培养法的一种变形,同时使用两个反应器,第一个反应器主要是为大量培养细胞,第二个反应器是为增加目的产物的含量。
P178一般连续培养的细胞产率高于分批培养的,但由于植物细胞生长速率慢,维持长时间的无菌状态是需要解决的基本技术问题。
第七章 生物反应器中的传质过程
P185反应液的流变学特性:是指液体在外加剪切力t作用下所产生的流变特性,简称流变特性。当给定的流体在外加剪切力的作用下,一定产生相应的剪切速率γ,两者之间的关系为该流体在给定温度和压力下的流变特性。
P186有多种经验方程来描述非牛顿流体的流变特性,其中最简单的形式是指数律方程。
P186指数律方程:
式中:K-稠密度指数,或称指数律系数Pa.s;n-流变性指数,或称指数律的方次。
对于牛顿型流体,n-1,K=μ。对于非牛顿型流体,将τ/γ定名为表观粘度。
P186一些稀薄的细菌发酵液;以水解糖或糖蜜为原料培养酵母的醪液;为噬菌体侵害的发酵液等均为牛顿流体。
P186丝状菌悬浮液菌呈丝状或团状。团状菌丝体是以稳定的球状积聚在一起而生长,其直径可达几mm。无论是丝状或团状,流变学特性都是非牛顿型流体。
P187一般丝状菌的发酵液呈假塑性流体、胀塑性流体等非牛顿性流体特性,并且发酵液的流动特性还随时间而变化。
P187流体特性因素都会对生化反应器内的质量与热量的传递、混合特性及菌体生长等产生影响。
P187流体力学研究方法包括数值模拟和实验研究。
P188根据Weisz的观点:"西勒准数为1,且无任何扩散限制时,细胞和其它成分的生物催化反应以最大反应速率而进行"。但事实上达不到。
P188反应系统中,反应物从反应液主体到生物催化剂表面的传递过程对生物反应过程影响很大,特别是基质的传质速率低于生物催化剂的反应速率时,生物催化剂的催化效率将受到基质传递速率的限制。因而,在一些发酵过程,如SCP和多糖发酵中,产物的生成速率可通过提高限制性基质的传递速率来加以改善。
P188在1m3培养基中每小时需要的氧是溶解量的750倍。因此,在生物反应过程中有效而经济地供氧是极为重要的。
P188微生物对氧的利用率首先取决于发酵液中氧的溶解度和氧传递速率。采取高密度培养方法提高生产效率时,高密度的细胞将使氧的消耗速度超过氧的传递速度。此时,氧的传递速度成为生物反应的限制性因素。
P189发酵过程中,有的微生物以菌丝团(或絮状物)的形式生长繁殖,为克服发酵过程中的扩散限制,可通过减小菌丝团尺寸的方法来解决。
P189微生物培养中的传质过程甚为复杂。几十年来,在提出的一些传质基本理论中, 被广泛用来解释传质机制和作为设计计算的主要依据的是双膜理论。
P189双膜理论的基本要点:1)在气液两个流体相间存在界面,界面两旁具有两层稳定的薄膜,即气膜和液膜。这两层稳定的薄膜在任何流体动力学条件下,均呈滞流状态。2)在气液界面上,两相的浓度总是相互平衡(空气中氧的浓度与溶解在液体中的氧的浓度处于平衡状态),即界面上不存在氧传递阻力。3)在两膜以外的气液两相的主流中,由于流体充分流动,氧的浓度基本上是均匀的,也就是无任何传质阻力,因此,氧由气相主体到液相主体所遇到阻力仅存在于两层滞流膜中。
P190根据双膜理论,对于氧的传递,以液相浓度为基准可得公式:
各字母的含义分别是:
kL为液膜传质系数;kG为气膜传质系数;Ci为气液界面上的平衡浓度;C为培养液主流中氧的浓度;C*为与气相氧分压相平衡的氧浓度;H为亨利常数;KL为以液膜为基准的总传质系数。
P191双膜理论中,各传质阻力的大小取决于气体的溶解度。如果气体在液相中的溶解度高,如氨气溶于水中时,液相的传质阻力相对于气相的可忽略不计;反之,对于溶解度小的气体,总传质系数KL,接近液膜传质系数kL,此时,总传质过程为液相中的传递过程所控制。
P191根据双膜理论得到的氧传质速率公式为:
Na--单位体积反应液中氧的传质速率mol/m3s;kLa--体积传质系数s-1;
P191营养物质通过细胞膜的传递形式主要有:被动传递(又称单纯扩散)、主动传递(又称主动运输)、促进传递(又称促进扩散)。
P191主动传递(又称主动运输):营养物从低浓度向高浓度的扩散,需消耗能量(代谢能)。
P191促进传递(又称促进扩散):营养物依靠载体分子(载体蛋白质或渗透酶)的作用而穿过细胞膜。
P192主动传递中,推动力是靠ATP水解释放的能量来进行的。
P192促进传递是借助载体分子完成的。促进传递的特征之一是其传递速率与酶促反应中的米氏方程类似。
P192亚硫酸盐法测定体积氧传递系数的方法原理:正常条件下,亚硫酸根离子的氧化反应非常快,远大于氧的溶解速度。当Na2SO3溶液的浓度在0.018~0.45mol内,温度在20-45℃时,反应速度几乎不变。所以,氧一旦溶解于Na2SO3溶液中立即被氧化,反应液中的溶解氧浓度为零。此时氧的溶解速度(氧传递速度)成为控制氧化反应速度的决定因素。过量的碘与反应剩余的Na2SO3反应,再用标准的Na2S2O3溶液滴定剩余的碘。根据Na2S2O3溶液消耗的体积数,可求出的Na2SO3浓度。将测得得反应液中残留的Na2SO3浓度与取样时间作图,由Na2S2O3消耗曲线的斜率求出dCNa2SO3/dt,再由公式求出kLa。
P192亚硫酸盐法的优点是适应kLa值较高时的测定,但对于大型反应器来讲,每次实验都要消耗大量的高纯度的亚硫酸盐。
P193由于亚硫酸盐法测定kLa是在非培养条件下进行的,因此所测kLa值与实际培养体系的kLa值存在差异。
P193利用氧电极进行kLa的测量有多种方法,动态法是常用方法之一。
P193动态法测定KLa的方法原理:通风培养液中氧的物料衡算:
当停止通风,有:
根据培养液中溶解氧浓度变化速率,可以求出QO2X。当液体的溶氧浓度下降到一定程度时(不低于临界溶解氧浓度)再恢复通气,培养液中溶解氧浓度将逐渐升高,最后恢复到原先的水平。
由 对C作图,从所得直线的斜率求出kLa值,并由截距得到C*。
对C作图,从所得直线的斜率求出kLa值,并由截距得到C*。
P194葡萄糖氧化法是在有氧条件下,利用葡萄糖氧化酶的催化作用,通过葡萄糖生成葡萄酸的反应,测定kLa的方法。
P195影响kLa的因素可分为三部分:操作变量、培养液的理化性质、反应器的结构三部分。
P195反应器中气泡流动方式分为两类:1)气泡自由上升;2)呈高湍流型(搅拌)。大型发酵罐归为前一类。
P195鼓泡式反应器的kLa中的a的大小取决于所设计的空气分布器、空气流动速率、反应器的体积、空气泡的直径等。
P195高湍流鼓泡式反应器,可利用下式估算kL:
P195鼓泡式反应器的kL关联式有:
P196低雷诺数条件下,气泡的运动服从Stokes 定律,气液间比表面积与气泡直径的三次方成反比,基于此,采用强烈搅拌操作,是为减小dB(气泡直径),从而增大a值。
P196气升式反应器中,通气量小时,空气通过小孔在液体中形成不连续的气泡。此时,气泡的大小可利用离开分布器的气泡所受的平衡力来确定。公式为:
P196机械搅拌罐中,气泡直径的计算公式为:
P197机械搅拌式反应器中的体积溶氧系数的概括公式为:
N—为搅拌器转速;K—有因次的系数,其随搅拌器形式、反应器的形状而变化;a、β和γ-经验指数。
其取值范围分别为α=0.3~1.2;β=0.2~1.0和γ=0.5~2.6。当α=0.4,β=0.5,γ=0.5时,上式称为Richard公式。
P198 Oconner的研究表明,常温下利用活性污泥法处理废水时,提高温度可增加kLa值。
P198在嗜热脂肪芽孢杆菌的培养过程中发现kLa与温度成正比,与μL成反比。
P198Richards方程把液体的粘度、密度、表面张力和气体溶质在液相中的扩散系数等都归入总的常数项内,不再作为参变量存在于关系式中。
P198反应液中的有机物和盐类等都会影响KLa。
P199发酵工业中,气泡平均大小的变化依赖于液体成分、气体的空塔速度和液体状态,是否湍流等。少量的盐或酒精加入到反应液中,会相应减少气泡的大小。
P199通用式发酵罐中搅拌器的组数及搅拌器之间的最适距离对溶氧有一定的影响。
P199一般地讲,当高径比为2.5时用多组搅拌器可提高溶氧系数10%,当高径比为4时采用较大空气流速和较大功率时,多组搅拌可提高溶氧系数25%。
P200带有搅拌装置的反应器都应安装适当的挡板,挡板可以使液体形成某种轴向运动,减少回旋运动,不让大量空气通过旋涡外逸,从而提高了气液混合效果,改善氧的传递条件。一般反应器可装4块挡板。
P202一般认为,发酵系统中氧由气相到液相的总传质系数近似等于液膜传质系数。
P203当氧的传递速率大于氧的消耗速率时,菌体的耗氧速率成为限制性因素。
P205菌丝团(菌丝球)中氧的传递模型简化后的公式为:
P205菌丝团的氧传递模型公式中的a反映了菌丝团中最大反应速率与最大传递速率之比。反应速率越大,传递速率越小,菌丝团内部缺氧就越严重,效率因子也就越低。
P207评价通风生化反应器的两个重要指标:kLa值和单位溶解氧的能耗(Np)。
P207提高氧传递速率Na的两条途径:一是提高氧传质推动力(C*-C);二是提高kLa值。提高氧传递速率的同时,应尽量减小通风搅拌功率的增加,以保证有较低的Np值。
第八章
P210生物反应器与化学反应器在使用中的主要不同点是生物(酶除外)反应都以“自催化"(Autocalalysis)方式进行,即在目的产物生成的过程中生物自身要生长繁殖。
p211生物反应器的分类(1)根据生物催化剂分类:酶反应器和细胞反应器;(2)根据底物加入方式分类:分批式、连续式、流加式(3)根据流体流动或混合状况分类:连续操作搅拌式反应器、连续操作活塞式反应器 。
p212生物反应速率主要指细胞生长速率、基质消耗速率和产物生成速率
P213生物反应器选型与设计的要点:1.选择适宜的生物催化剂。这包括要了解产物在生物反应的哪一阶段大量生成.适宜的pH和温度,是否好氧和易受杂菌污染等。2.确定适宜的反应器形式。3.确定反应器规模.几何尺寸.操作变量等。4.传热面积的计算。5.通风与搅拌装置的设计计算。6.材料的选择与确保无菌操作的设计。7.检验与控制装置。8.安全性。9.经济性。
P214生物反应器中的热量平衡可表示为:
式中Qmet为微生物代谢或酶活力造成的单位体积产热速率;Qag为搅拌造成的单位体积产热速率;Qgas为通风造成的单位体积产热速率;Qacc为体系中单位体积的积累产热速率;Qexch为单位体积反应液向周围环境或冷却器转移热的速率;Qevap为蒸发造成的单位体积热损失速率;Qsen为热流(流出一流入)造成的单位体积敏感焓上升的速率。
P214实际生物反应过程中的热量计算,可采用如下方法:1.通过反应中冷却水带走的热量进行计算。2.通过反应液的温升进行计算。3.通过生物合成进行计算。4.通过燃烧热进行计算。
P215单相系统酶反应器有:搅拌罐、超滤膜反应器;多相系统酶反应器:通用式发酵罐、固定床或填充床、流化床、膜式反应器、悬浊气泡塔。
P215从几何形状或结构来划分,酶反应器大致可分为罐型、管型和膜型三类。
P216理想型连续式操作的反应器分为连续操作活塞式反应器(CPFR)和连续操作搅拌式反应器(CSTR)。
P216决定酶反应器设计和操作性能的参数有停留时间、转化率、反应器的产率、酶的用量、反应器温度、pH值和底物浓度等。当副反应不可忽视时,选择性Sp也是很重要的参数。
P216停留时间t是指反应物料进入反应器时算起,至离开反应器时为止所经历的时间。
P216如果反应器的容积为V,物料流入反应器中的体积流量为F,平均停留时间τ的定义式为: ;τ又称空时(空间时间space time),其倒数1/τ称为空速(空间速度space velocity)。
;τ又称空时(空间时间space time),其倒数1/τ称为空速(空间速度space velocity)。
P217转化率X(也称转化分数conversion or fractional conversion)是表明供给反应的底物发生转变的分量。
分批式操作中,底物的初始浓度为[S]0,反应时间t时的底物浓度为[S]t,此时,底物[S]的转化率为:
连续式操作中,流入反应器内的底物浓度为[S]in流出液中底物的浓度为[S]out,此时转化率:
P217反应器生产能力Pr(productivity)的定义是单位时间、单位反应器体积内生产的产物量。
分批式操作中, 式中Pt为时间t时单位反应液体积中产物的生成量。
式中Pt为时间t时单位反应液体积中产物的生成量。
连续式操作中, 式中Pout为单位体积流出液中的产物量。
式中Pout为单位体积流出液中的产物量。
P217选择性Sp(selectivity)是在有副反应发生的复合反应中,能够转变为目的产物的底物变化总量中,实际上转变为目的产物的比率。
P217由底物S生成目的产物P的选择性[S]p为: [S]p表明了整个反应的平均选择性。式中asp是指从1mol底物S中所得到产物P的摩尔数,是由反应的计量关系而决定。
[S]p表明了整个反应的平均选择性。式中asp是指从1mol底物S中所得到产物P的摩尔数,是由反应的计量关系而决定。
P218 CPFR型酶反应器也称为活塞流式反应器或平推流式反应器。CPFR具备以下特点:在正常的连续稳态操作情况下,在反应器的各个截面上,物料浓度不随时间而变化;反应器内轴向各处的浓度彼此不相等,反应速率随空间位置而变化;由于径向有严格均匀的速度分布,即径向不存在浓度分布,故反应速率随空间位置的变化只限于轴向。
P220 CSTR型与CPFR型反应器性能的比较中,在相同的工艺条件下进行同一反应,达到相同转化率时,两者所需的停留时间不同,CSTR型的比CPFR型反应器的要长,也就是前者所需的反应器体积比后者大。
P220 CSTR型与CPFR型反应器性能的比较中,在相同的工艺条件下进行同一反应,转化率越高,CSTR中所需酶的相对量也就越大。
P222 CSTR型与CPFR型反应器性能的比较中,0级反应时,CSTR与CPFR内酶活力的衰退没有什么区别。一级反应时候,CPFR产量的下降要比CSTR快得多,因而CPFR中酶的失活比CSTR中更为敏感。
P222 CSTR型与CPFR型反应器性能的比较中,在CPFR中,虽然出口端浓度较低,但在进口端,底物浓度较高;CSTR中底物总处于低浓度范围。如果酶促反应速率与底物的浓度成正比,那么对于CSTR而言,由于整个反应器处于低反应速率条件下,所以其生产能力也低。
P226 机械搅拌式发酵罐,是指既具有机械搅拌又有压缩空气分布装置的发酵罐。
P229 机械搅拌罐的混合主要是通过机械搅拌来实现。机械搅拌不仅可促使培养基混合均匀,而且有利于增加气液接触面积,提高溶氧速率。
P229 生物反应器中常使的搅拌器型式有:螺旋桨、平桨、涡轮桨、自吸式搅拌桨和栅状搅拌桨等。另外,翼型桨也已开始广泛应用于发酵生产,并取得较好效果。
P230 不通风情况下,牛顿型流体通过因次分析可得搅拌功率计算公式: 具体为:
具体为:
式中:Np为功率准数,其物理意义为机械搅拌力与惯性力之比;ReM为搅拌雷诺准数,其物理意义为惯性力与粘滞力之比;FrM为搅拌弗鲁特准数,其物理意义是搅拌加速度与重力之比;K为与搅拌器形式、反应器几何尺寸有关的常数。
P231 不通风情况下,在全档板条件下,当R>104,液体处于湍流状态,搅拌功率计算公式为:
P232 通风时搅拌器的轴功率消耗降低,Michel等人提出了应用较广泛的通风时的搅拌功率Pg与工作变量间的经验公式:
P236 鼓泡式反应器的功率消耗可由下式计算:
P239 连续式通风固态发酵设备有塔式,转鼓式和回转式等多种形式。塔式通风固态发酵设备外形为塔式,内有两至六层塔板;转鼓式通风固态发酵设备外形为放倒的圆柱体;回转式固态发酵设备为一圆环形装置。
P246在生物反应器的设计与操作中,至少有两点必须明确:1.目的反应是如何进行的,即所期望的生化反应是通过何种渠道完成的。2.生物化学反应中哪些反应的反应速度快、哪些反应的反应速度慢。
P246生物反应器的放大方法可分为(1)数学模拟放大;(2)因次分析法放大;(3)经验法则放大(包括反复实验法、部分解析法放大等)。
第九章
P252为提高工程菌株发酵效率,人们最初是建立目的产物的产率与相关基因表达能力、宿主的遗传特性及发酵过程中操作条件等之间定性与定量的关系。
P252当基因的表达仅取决于细胞所含基因拷贝数时,工程菌细胞中所含质粒的稳定性及其复制与表达的能力成为关键因素。这样便简化了质粒的复制与表 达的动力学描述。
P256一般认为,工程菌株的质粒不稳定过程分为两个阶段,其一是工程菌株细胞分裂时,某子细胞未能得到质粒或质粒缺失等;其二是由于质粒保持菌株与失去质粒菌株的生长速率的差异,后者的生长速率可能更快一些,致使后者所占总菌数的比例越来越大。
P257目前,代谢工程技术已在许多领域得到应用,如异源蛋白的生产;扩大底物利用范围(特别是可再生资源的利用);生产新物质;环境污染物的降解;提高菌体对环境的适应能力;阻断或降低副产物的生成;代谢产品生产速率和生产能力的提高;植物代 谢工程;动物代谢工程;人体和组织代谢工程--新药发现及人类疾病诊断和基因治疗。
P257代谢工程是利用DNA重组技术对特定的生化反应进行修饰或引入新的反应以定向改进产物的生成或细胞的性质的学科。
P257代谢工程包括分析与合成两个步骤。
P258代谢工程包含代谢途径、代谢通量和在简单途径及更实际的代谢网络中控制代谢通量的因素等概念。
p258代谢工程的原理是在对细胞内代谢途径网络系统分析的基础上进行有目的的改变,以更好地利用细胞代谢进行化学转化、能量转导合成、分子组装,它的研究对象是代谢网络,依据代谢网络进行代谢通量分析(metabolic flux analysis, MFA)和代谢控制分析(metabolic control analysis, MCA),并检测出速率控制步骤,最终的目的是改变代谢流,提高目的产物的产率。
P261代谢通量分析(MFA)是用计量矩阵模型表示胞内的主要反应。根据代谢途径中各反应的计量关系及实验中测得的底物的消耗速率、产物的生成速率及细胞的生成速率确定代谢网络的通量分布图。
P261同位素标记实验的基本原理是使用同位素标记的底物进行微生物培养,通过测量胞外通量及胞内标记信息,对胞内通量求解。
P265系统生物学是系统性地研究一个生物系统中所有组成成分 (基因、mRNA、 蛋白质等)的构成,以及在特定条件下这些组分间的相互关系,并分析生物系统在一定时间内的动力学过程。
P266系统生物学的研究平台:1基因组学2转录组学3蛋白质组学4代谢组学5相互作用组学
P274合成生物学的学科任务主要有两个方面:①进行新的生物零件、组件和非天然控制的细胞活动的分子网络系统的设计与构建,从而创建全新的完整生物系统乃至人工生命体;②对现有的、天然存在的生物系统的重新设计与改造,使其能按照需要完成特定的生物学目标。简单地说,合成生物学就是通过人工设计和构建自然界中不存在的生物系统来解决能源、材料、健康和环境等问题。
P274合成生物学强调“设计"和“重设计"。
P274合成生物学的基本工艺流程分为设计、建模、合成和分析4个步骤。
P274“工程化”是合成生物学的一个显著特点,也是合成生物学区别于现有生物学其他学科的主要特点。
P274合成生物学的研究目标非常明确:从头合成新的生命体,或者对已有生命体进行工程化改造,从而实现新的功能。
P275目前,合成生物学的研究主要集中在以下几个方面:
①生物元件标准化及生物模块的设计与构建;
②底盘基因组的设计和构建;
③基因组的设计、合成与组装;
④基因线路和代谢网络的设计与构建;
⑥工程生物系统的计算机模拟。
第三章 酶促反应
P22确保酶活力稳定的稳定化方法及原因 p28米氏方程3假说 p29求km和rp,max方法(Lineweaver Burk法、Eadie-Hofstee法) p46 DA准数公式及意义 p47内外扩散过程的对比 p54不同温度对酶促反应的影响
第四章 微生物反应动力学
P100微生物生长动力学的数学模型,根据理论分类 P101Monod方程
第五章 反应器操作
p121分批培养过程的状态方程式 P123反复分批式培养示意图及衡算式 p130单级CSTR培养系统中菌体、限制性基质及产物的物料衡算数学表达式及各字母含义
第七章 传质
P190双膜理论 P190根据双膜理论得到的公式 P192KLa的测定方法(亚硫酸盐法、动态法) P197机械搅拌式反应器中的体积溶氧系数的概括公式、Richard公式
第八章
p213生物反应器选型和设计的要点 P214生物反应器中热量平衡表示公式及字母含义 P218 CPFR型酶反应器特点
计算题
第三章 酶促反应
p27 一级反应计算
题目:卵状假单胞菌在分批式反应中先将葡萄糖转变为葡萄糖内酯,再转化为葡萄糖酸。此反应为一级反应,反应式如下:

求中间产物(葡萄糖内酯)浓度达最大时所需的时间tmax及其最大浓度值[L]max。已知反应常数k1和k2分别为0.87h-1和0.7h-1。[G]0(初始葡萄糖浓度)=38mg/ml。(e取2.71828)
解:当葡萄糖内酯浓度达到最大值时,此时d[L]/dt=0,所以
=(ln 0.87/0.7)/(0.87-0.7)=1.28h
=(0.87×38)/(0.7-0.87)×(e-0.87x1.28-e-0.7x1.28)=15.5g/L
即当tmax=1.28h时,葡萄糖内酯浓度达到最大值15.5g/L。
第四章 微生物反应动力学
P87 物质恒算计算 例4-1求系数 例4-2呼吸商 例4-3酵母经验分子式 P91 P93得率计算 例4-4 例4-5 Yx/s Yx/o P100例4-7求td P115 课后19 求系数和Yx/s Yx/o
第五章 微生物反应器操作
P123 例题5-3 初始基质浓度 F 取出率
第七章 传质
P194 最大细胞浓度
题目:采用100L通用式发酵罐培养大肠杆菌,通风比(每分钟单位体积培养液中的通风量,下同)0.8L/(L▪min),kLa=0.0417/s,求确保满足Oo2=8.89×10-5g/(g.s)(以氧/细胞计),ccri=0.2mg/L,c*=7.3mg/L时,不出现缺氧状态,求此状态下的最大细胞浓度。
解:为确保不出现缺氧,应满足供氧速率-耗氧速率,即

所以,在此条件下最大细胞浓度为

=0.0417(7.3×10-3-0.2×10-3)/8.89×10-5=3.33(g/L)
2022A 米氏方程三点假说 Monod方程并解释各字母的含义 计算反应中得率系数Yx/s和Yx/o
2022B 亚硫酸盐法测定体积氧传递系数的原理 影响固定化酶促反应的主要因素 求Yx/s和Yx/o及Yc
2023A 双膜理论基本要点 体积传质系数的测定中动态法的测定方法 求出酵母的经验分子式
2023B 双膜理论的推导公式 DA准数的物理意义 计算反应式中的计量系数
2024A CPFR型酶反应器 .Lineweaver Burk法 卵状假单胞菌在分批式反应
2024B 生物反应器选型与设计的要点 反复分批操作示意图写出菌体量的衡算、产物浓度的衡算 采用100L通用式发酵罐培养大肠杆菌,求最大细胞浓度
2024AA 米氏方程的三点假设中第三点、方程式、各个字母的含义 计算初始基质浓度
2024BB 微生物生长动力学的数学模型分类内容 求倍增时间